Anatomy of the Human Eye



|
Anatomy of the Human Eye |
|
The eyes, the "sense of visual perception", rank among the most important sensory organs of the human organism. They provide us with a constantly updated view of the environment. The following explanations refer to the right eye. The eyeball or globe (Latin bulbus oculi, briefly bulbus, ø approx. 24 mm, nearly spherical [Pschyrembel, 1994]) lies protected in the eye socket (orbita), that forms a cavity in the cranium. The globe is onion skin-like composed of three layers [Schäffler and Schmidt, 1998]:
•sclera (leather skin): outer eye skin,
•choroidea (vein skin): middle eye skin,
•retina: inner eye skin.
The six extraocular eye muscles are responsible for the movement of the globe. The four straight eye muscles (musculi recti) and the two oblique eye muscles (musculi obliqui) originate in the orbita and insert at the sclera of the globe. The figure shows the muscle origins in the orbita and their insertion on the globe. Each eye muscle affects the globe in three components, whereas the muscle path determines the main direction of pull. The main effect of each muscle can be derived from its name [Brugger, 2000].
•superior rectus muscle (upper straight eye muscle): upward,
•inferior rectus muscle (lower straight eye muscle): downward,
•lateral rectus muscle (outer straight eye muscle): sideways outward,
•medial rectus muscle (inner straight eye muscle): sideways inward,
•superior oblique muscle (upper oblique eye muscle): downward and incyclorotation,
•inferior oblique muscle (lower oblique eye muscle): upward and excyclorotation.
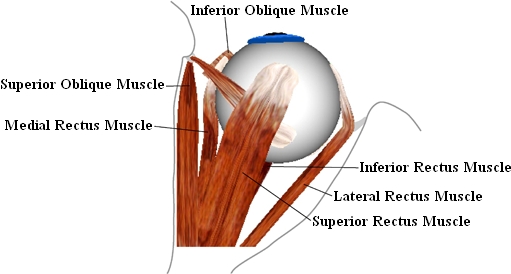
Top View of the Right Eye
The musculi recti originate at the posterior end of the orbita and are arranged in pairs right-angled to each other. Their end tendons join at the anatomic origin to a circular tendon plate (Zinn's ring) and their insertions lie before the equatorial plane of the globe (cf. [Günther, 1986]). In contrast, the musculi obliqui insert behind the globe equator and pull diagonally forward. The superior oblique muscle is the longest of all eye muscles. Starting from its origin near Zinn's ring it is running above the globe towards the nasal frontal bone, its tendon is slipping through a cartilaginous hole (the trochlea) and from there proceeds directly to its insertion. The inferior oblique muscle originates from the nasal edge of the bony orbita, runs below the globe around the inferior rectus muscle and inserts at the back area of the globe. In the area of intersection between inferior oblique muscle and inferior rectus muscle, both muscles are connected by the ligamentum lockwood [Günther, 1986].
Each eye muscle consists, apart from the purely muscular portion, also of a tendon which connects the muscle at the origin on one side and at the point of insertion on the other side. The overall length (muscle and tendon) of the extraocular muscles is very different. Thereby the lengths of the tendons show the largest differences (cf. [Kaufmann, 1995]). The inferior oblique muscle has the shortest tendon with 0 to 2 mm and the superior oblique muscle has the longest tendon with 25 to 32 mm. The actual muscle length (without the tendon) lies between 30 mm (musculi obliqui) and 39 mm (inferior rectus muscle). Due to the insertion lying before or behind the equator of the globe, each muscle partially contacts the globe's surface. At the point of tangency, the muscle loses contact to the globe and pulls toward its origin (cf. Fig.). With each movement of the globe, the relative position of a muscle's insertion changes with respect to the orbita.
If the muscles could move freely between insertion and origin during an eye movement (hypothesis of the string model), a shift of the muscle path on the globe surface would occur, especially in tertiary gaze positions. Thereby the muscle path and consequently the direction of pull would change considerably according to the current eye position (loss of the main direction of pull). In order to avoid this, connective tissue surrounds the globe and stabilizes the muscles within the area of the point of tangency. These stabilizers are called pulleys (cf. [Buchberger and Mayr, 2000], [Miller and Demer, 1996]).
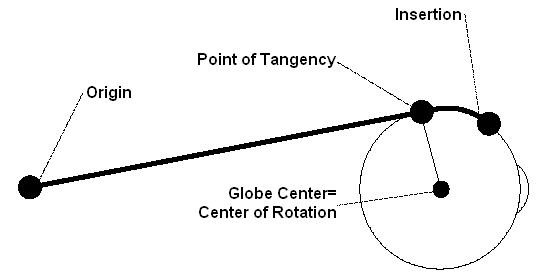
Schematic Illustration of a Straight Eye Muscle with Origin, Point of Tangency and Insertion
The movement of the globe approximately corresponds to a rotation of an object in the three-dimensional space around a certain axis. The globe center can be regarded as rotation center. The line of sight is a vector from the globe center through the center of the pupil. Perpendicular to this vector, the vertical and the horizontal axes are defined, whereas the intersection of these three axes lies in the globe center (cf. Fig.). Eye positions can be classified by their rotational properties (cf. [Kaufmann, 1995]):
•Primary position: The eye looks straightforward, with the head fixed and upright. It is assumed, that in this position all muscles exhibit minimum force (rest tonus). From this position all the other gaze positions can be reached with little energy demand.
•Secondary position: Starting from the primary position, a rotation around the horizontal or vertical axis is performed (Fig. b). The eye looks to the left or to the right or respectively upwards or downwards.
•Tertiary position: Starting from the primary position, a rotation around the horizontal and vertical axis is performed (Fig. c). The eye looks e.g. to the left and up or to the right and down. The combination around two axes can also be represented by one single rotation axis, which lies in the plane spanned by the horizontal and vertical axis (Fig. d).
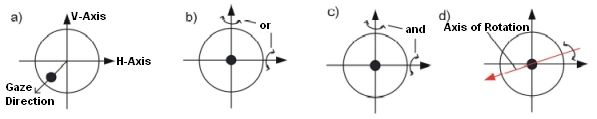
Line of Sight, Vertical and Horizontal Axis; Rotations to Other Eye Positions
Both eyes can only be moved together in binocular community, which means that the movement of only one eye is normally impossible (cf. [Günther, 1986]). Eye muscles are able to reposition the eye accurately and very fast, moreover, they can hold a certain position without exhaustion.
The rotation of an eye around a certain axis results in a certain gaze position and thus also realigns the line of sight to a new gaze direction. The gaze direction designates the orientation of the eye, whereas the gaze position should always come along with a fixation of the eye onto an object.
As mentioned before, each muscle is defined through its origin, point of tangency and insertion. The muscle paths from the origin to the insertion on the globe are, however, additionally influenced by connective tissue, the so-called "pulleys".
The following figure shows a schematic illustration of these anatomical structures:
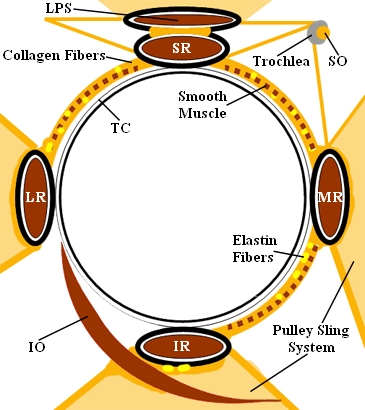
Representation of the Orbital Connective Tissue [Miller and Demer, 1999]
A "pulley" encloses a muscle as a circular structure and is connected with the orbital wall (retinacula) and other connective tissue structures (inter-muscular membrane). Smooth musculature and nerves are contained in this apparatus. Tendons and muscles travel through these pulleys, whereas the pulleys themselves are fixed relative to the orbita and are located near the bulbus equator in primary position
Anatomical Influence of Pulleys on Eye Movements
Before the discovery of pulleys as functional elements (1995) all models were based on the assumption that the anatomical origin of an eye muscle is also the functional origin. This again leads to the assumption, that the rotation axis, around which a muscle rotates the globe, is perpendicular to the plane defined by a muscle's point of tangency, center of rotation and anatomical origin. With the discovery of pulleys as functional elements, this definition of the axis of rotation is no longer valid. Therefore, according to the pulley model, the effective pulley location can be assumed as the actual functional origin of an extraocular eye muscle, since the muscle changes its main direction of pull at this position due to the presence of the pulley. As a consequence, the axis of rotation now has to be defined through a muscle's point of tangency, the center of rotation and a muscle's effective pulley location. The string model of Robinson (1975) as well as the so-called "intensified string model" according to Kusel and Haase (1977) have already tried to consider the influence of pulleys via a component which reduces the sideslip angle for each muscle. In contrast, there are newer models like Orbit™, Eyelab and SEE-KID, which already include pulleys as anatomical components and as a consequence also provide more accurate results compared to clinical data.
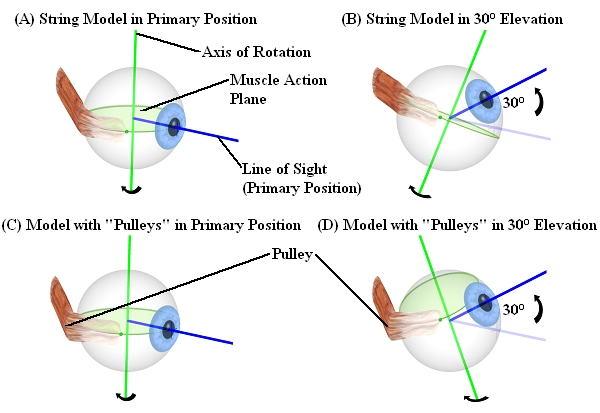
Difference Between Conventional Models (Above) and Models with Pulleys (Below)
The discovery of pulleys as functional elements explained, why all other models were less successful in comparison with real clinical data. Also the implementation of surgery simulations had to be adapted to these new findings.
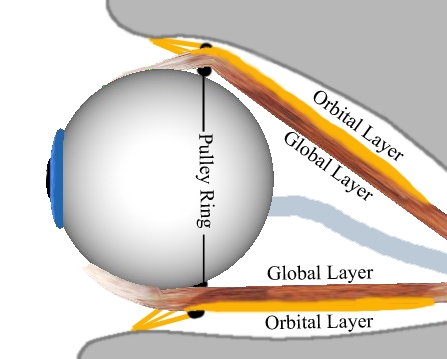
Global Layer and Orbital Layer of Superior Rectus Muscle and Inferior Rectus Muscle
Previous studies with mammals have already suggested that the extraocular eye muscles consist of two different layers. As shown in the preceding figure, the global layer runs continuously from the origin (near Zinn's ring) to the muscle's insertion on the globe. In contrast, the orbital layer ends slightly behind the insertion.
Several studies by [Demer et. al., 2000] and [Clark et. al., 2000] have shown, that each of these two layers has a different function. While the global layer is primarily responsible for the movement of the eye into a certain gaze position and runs up to the insertion, the orbital layer ends in the area of the pulleys and actively moves the pulleys when the eye is looking in a secondary or tertiary position. It could be shown by various studies, that the pulley position moves backwards during the contraction of an eye muscle and forwards during relaxation. These findings led to the formulation of the "Active-Pulley-Hypothesis", which is implemented in the SEE++ system in form of the active-pulley model. SEE++ is the first simulation system implementing the "Active-Pulley-Hypothesis".